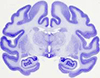
Nissl, acetylcholinesterase, cytochrome oxidase, myelin, parvalbumin, calbindin, and Cat 301 preparations allowed the ventral anterior and ventral lateral thalamic regions, parvocellular and magnocellular subdivisions of ventral anterior nucleus, and anterior and posterior subdivisions of ventral lateral nucleus of monkeys to be identified. 

Magnetic resonance imaging showed low intensity areas (left posterior limb of internal capsule, left cerebral peduncle of middle brain, a part of left substantia nigra, left amygdala, ventral posterior lateral nucleus and ventral anterior nucleus of left thalamus, left lateral geniculate body, and left occipital lobe) in T1 weighted image, due to the infarct in the left anterior choroidal artery territory.
In the course of mapping the dorsal thalamus, we also studied neurons in a subset of thalamic nuclei (the caudal part of the ventral lateral nucleus (VLc), the oral part of the ventral posterior lateral nucleus (VPLo), the parvocellular part of the ventral anterior nucleus (VApc)) lateral to the MD and just across the internal medullary lamina.
Statistically significant, 40% decrease (P=0.055) of Bmax for [ (3)H]muscimol was observed in the ventral anterior nucleus pars densicellularis (VAdc, the main pallidal projection territory in the thalamus) 1 week after globus pallidus lesioning and a 36% decrease (P=0.017) 4 months post-lesioning.
The labeled cerebellar terminals were distributed throughout the posterior VL (VLp), whereas the labeled pallidothalamic terminals were concentrated in the anterior VL and the ventral anterior nucleus.
The ventral anterior nucleus (VA) and the ventral lateral nucleus pars oralis (VLo) contained a greater density of pallidal labeling while a greater density of cerebellar label was observed more caudally in the ventral posterior lateral nucleus pars oralis (VPLo) as well as in nucleus X (X).
In whatever part of the substantia nigra the injection was made, numerous axonal endings were found to be distributed within different thalamic regions: the ventral anterior nucleus and mainly its magnocellular part, the most ventromedial part of the ventral lateral nucleus, and the mediodorsal and parafascicular nuclei.
Other branches ascended to the thalamus and broke into 10-15 thinner collaterals that ran through most of the ventral anterior nucleus, where they terminated as typical plexuses.
Computed tomography and magnetic resonance imaging revealed cerebral haemorrhage in the right thalamus involving the ventral anterior nucleus, medioventral nucleus, mamillothalamic tract, internal medullary lamina, and mediodorsal nucleus.
Lesion location data demonstrated that the ventral anterior nucleus of the thalamus was always spared; the ventral posterior (lateral and medial) nucleus was always damaged, and the ventral lateral nucleus was frequently damaged.
To better understand the contribution of cerebellar- and basal ganglia-receiving areas of the thalamus [ ventral posterolateral nucleus, pars oralis (VPLo), area X, ventral lateral nucleus, pars oralis (VLo), or ventral anterior nucleus, pars parvicellularis (VApc)] to movements based on external versus internal cues, we temporarily inactivated these individual nuclei in two monkeys trained to make visually triggered (VT) and internally generated (IG) limb movements.
In the lateral thalamus, cells exhibiting random or rhythmic low-threshold calcium spike bursts were found preponderantly in the ventral anterior nucleus (53.4%) and in the ventral lateral anterior nucleus (52.7%).
The cerebellar nuclei primarily projected to posterior (VLp), medial (VLx), and dorsal (VLd) divisions of the ventral lateral nucleus; the pallidum largely projected to the anterior division (VLa) of the ventral lateral nucleus and the parvocellular part of the ventral anterior nucleus (VApc).
Seven days post-lesion, a 20-30% decrease in Bmax values (P < 0.05) was found in the nuclei receiving input from the lesioned nucleus reticularis thalami sector (the mediodorsal nucleus and densicellular and magnocellular parts of the ventral anterior nucleus) without changes in affinity.
The proportion of cells in each category was found to vary between each of the cerebellar-receiving [ oral portion of the ventral posterolateral nucleus (VPLo) and area X] and basal-ganglia-receiving [ oral portion of the ventral lateral nucleus (VLo) and parvocellular portion of the ventral anterior nucleus (VApc)] nuclei that were examined.
Each injection resulted in anterograde labeling in the three subdivisions of the ventral anterior nucleus (pars parvicellularis, VApc; pars densicellularis, VAdc; and pars magnocellularis, VAmc) and in the ventral lateral nucleus (VL).
We found that the SMA thalamocortical neurons occupied a wide band extending from the ventral anterior nucleus pars principalis (VApc) through the ventral lateral nucleus pars oralis (VLo) and the ventral lateral nucleus pars medialis (VLm) and into to the ventral lateral nucleus pars caudalis (VLc) including a portion of ventral posterior lateral nucleus pars oralis (VPLo) and nucleus X.
Overlapping thalamic territories between the thalamostriatal neurones projecting to areas of the caudate nucleus and the nigrothalamic connections were observed in the rostral nuclei of the central thalamic group (ventral anterior nucleus, ventral anterior-ventral lateral complex and ventral medial nucleus) and, more restricted, in the rostral (rhomboid, paracentral, ventral lateral, dorsal mediodorsal nuclei) and caudal intralaminar nuclei (centromedian-parafascicular complex).
Overlapping thalamic territories between the thalamostriatal neurones projecting to areas of the caudate nucleus and the nigrothalamic connections were observed in the rostral nuclei of the central thalamic group (ventral anterior nucleus, ventral anterior-ventral lateral complex and ventral medial nucleus) and, more restricted, in the rostral (rhomboid, paracentral, ventral lateral, dorsal mediodorsal nuclei) and caudal intralaminar nuclei (centromedian-parafascicular complex).
The neurons that project to the FEFsem are distributed in (1) the rostral portion of the ventral lateral nucleus, pars caudalis, (2) the caudal portion of the ventral lateral nucleus, pars caudalis, (3) the mediodorsal nucleus, (4) the ventral anterior nucleus, pars parvocellularis, and (5) the ventral anterior nucleus, pars magnocellularis.
In the present study, we investigated whether a dorsal thalamic region comparable to the motor part of the mammalian ventral tier (the ventral anterior nucleus, the ventral lateral nucleus, and the oral ventroposterolateral nucleus) exists in pigeon.
After those injections, dense plexuses of anterogradely labelled varicosities were found in common thalamic nuclei, including the parvocellular ventral anterior nucleus (VApc), the dorsal VL (VLd), and the rostrodorsal part of the parafascicular nucleus (PF).
We found dense single label in the central portion of the ventral anterior nucleus pars principalis (VApc) and the ventral lateral nucleus pars oralis (VLo) following the GPi injections or in the central portion of the ventral posterior lateral nucleus pars oralis (VPLo) and nucleus X (X) following the cerebellar nuclei injections.
In addition, the medial region of the ventral striatum receives numerous projections from the central superior lateral nucleus, the magnocellular subdivision of the ventral anterior nucleus, and parts of the mediodorsal nucleus.
To our knowledge, only one such case has been reported after a left thalamic lesion, located in the ventral anterior nucleus, the mamillo-thalamic tractus and the genu of the internal capsule.
The most medial band included area 6-projecting neurons in the anterior nuclei, the rhomboid nucleus, the ventral anterior nucleus (VA), ventromedial nucleus (VM) and mediodorsal nucleus (MD).
Following tritiated amino acid injections into the substantia nigra pars reticulata (SNr) and WGA-HRP injections into the contralateral cerebellar nuclei, we found that the nigrothalamic and cerebellothalamic afferents distribute to three main targets: the central portion of the ventral anterior nucleus (VA) and the ventral lateral nucleus (VL), the internal medullary lamina (IML) region, which includes the paralaminar VA, the mediodorsal nucleus (MD) and the central lateral nucleus (CL), and finally the ventromedial nucleus (VM).
The damage implicated the ventral anterior nucleus, the mamillothalamic tract, and the rostroventral internal medullary lamina.
The nigrothalamic label occupied a longitudinal band extending from rostral ventral anterior nucleus (VA) through to caudal mediodorsal nucleus (MD).
The habenula and the ventral anterior nucleus were among the most weakly innervated nuclei.
These PR neurons were located in rostral portions of VLc, in the oral part of the ventral lateral nucleus (VLo), and in the parvicellular part of the ventral anterior nucleus (VApc).
An EM-autoradiographic technique was used to identify the ultrastructural features and synaptic sites of nigral afferents to the ventral anterior nucleus pars magnocellularis (VAmc) of the rhesus monkey thalamus.
There were also smaller changes in portions of the thalamus (ventral anterior nucleus, parafascicular nucleus) and premotor cortex.
Six rat brain structures (external globus pallidus, ventral anterior nucleus of thalamus, entopeduncular nucleus, substantia nigra pars reticulata and superior colliculus) presented an increase of 2-DG labelling after acute injection of this toxic compound.
In the ventral anterior nucleus, GABA-positive neuronal profiles formed typical clusters, whereas they were more uniformly distributed in the posterior nuclei of the ventral group.
Thalamostriatal projections from the ventral anterior nucleus (VA) were mapped by using autoradiographic and horseradish peroxidase techniques in the dog.
A considerable number of neurons in the ventral anterior nucleus (VA) send axons to areas 4 and 6.
A prominent projection to area 5a arises from the posterior (Po) and ventral lateral (VL) complexes; less substantial projections originate in the ventral anterior nucleus (VA), the lateral intermediate complex (LI), and the central lateral nucleus (CL).
Thus, cells in the rostromedial part of the ventral anterior nucleus project to the head of CD and some cells in the rostral part of the ventromedial nucleus project to the head of CD and to PU.
In cases with injections placed in the lateral part of area 4, dense accumulations of label were present in the lateral part of the ventral anterior nucleus (VA), the central part of the ventral lateral nucleus (VL), the ventral half of the ventral posterior inferior nucleus (VPI), the caudal part of the central lateral nucleus (CL), and the centrum medianum (CM).
Diencephalic section of the brain at the level of the ventral anterior nucleus weakened but did not entirely eliminate rhythmic afterdischarge induced by stimulation of the claustrum in the anterior parts of the neocortex.
An injection of the MI forepaw area localized to the rostral and central part of the posterior cruciate gyrus resulted in a band of labeled neurons occupying the dorsal extent of VL and continuing into the ventrolateral aspect of the ventral anterior nucleus (VA).
Rather there are longitudinally oriented strips of terminal labeling which extend through all divisions of the ventral lateral nucleus, i.e., the VLps, the VLc, the VLo, as well as nucleus X, the oral division of the ventral posterolateral nucleus (VPLo), the central lateral nucleus (CL), and the most caudal region of the ventral anterior nucleus (VA).
Such targets include the visceral cell columns of the oculomotor complex, the rostral interstitial nucleus of the medial longitudinal fasciculus, and the magnocellular division of the ventral anterior nucleus.
Within the thalamus, projections to the ventral lateral nucleus were scanty compared to those in the ventral anterior nucleus.
The latencies of the potentials, similarity in shape and in depth of reversion suggest that the cerebello-cortical responses in the pericruciate cortex are mediated through the thalamic ventro-lateral nucleus making synapses in III-IV cortical layers while responses in the parietal cortex--through the thalamic ventral anterior nucleus making synapses in more superficial layers..
In autoradiographic studies, EPN axons were found to terminate in a J-shpaed region in the dorsal and medial part of the ventral anterior nucleus (VA) and the rostral portion of the adjacent ventral lateral nucleus (VL).
Each population of cells, projecting to the ventral anterior nucleus of the thalamus (VA), centrum medianum (CM), the lateral habenula (LHB), and the pedunculopontine nucleus (PP), was distributed throughout the nucleus.
An accumulation of radioactivity was found in the ventral medial nucleus and in the ventromedial part of the ventral anterior nucleus.
Lesions of the ventral anterior nucleus of the thalamus only impaired the feeding response to glucoprivation.
Following large dorsomedial lesions, small numbers of degenerating fibers were traced through the medial part of the magnocellular ventral anterior nucleus (VAmc) and the ventral part of the internal medullary lamina surrounding the anterior nuclei.
Rostrolateral nigral neurons transported radioactive label preferentially and abundantly to thalamic nuclei; localized isotope was found in parts of three thalamic nuclei, the medial part of the ventral lateral nucleus (VLm), the magnocellular part of the ventral anterior nucleus (VAmc), and the paralaminar part of the dorsomedial nucleus (DMpl)9 Lateral neurons in the caudal half of the SN transmitted radioactive label to the same thalamic nuclei as rostrolateral nigral neuron.
-
[ View All ]